更新时间:2017-01-19 09:01 浏览: 次 作者:张迪 文章来源: 生物通

Francisco Mojica不是第一个发现CRISPR的,但他可能是第一个为它神魂颠倒的科学家。他记得1992年的那一天,当他第一次看到这个即将开启生物技术革命的微生物免疫系统。当时他正在审视嗜盐微生物Haloferax mediterranei的基因组序列数据,并注意到14个不寻常的DNA序列,每个30个碱基长。这些序列大致相同,正向和和反向,每35碱基左右重复一次。很快,他发现了更多这样的序列。Mojica为此着迷,并以此作为他在西班牙阿利坎特大学的一个研究焦点。
这是一个不一般的决定,Mojica实验室多年来缺乏资金。在各种会议上,Mojica会“逮住”每个可以找到的重要人物,询问他们对这些奇怪的小重复序列的看法。但他们警告Mojica说,“不要太在乎这些重复。在许多生物体中有很多重复,很多年前我们就已经知道了,但仍不清楚它们中有多少是有用的。”
如今,科学家们越来越了解这种被命名为CRISPR(成簇规律性间隔的短回文重复序列) 的元件,并且将其作为了一种研究工具,然而虽然大多数生物医学的研究人员更关注的是CRISPR-Cas9如何用于编辑基因,但是Mojica和其他微生物学家仍然 希望了解有关这个系统的一些基本问题及工作原理:它是如何演变的?如何影响微生物进化的?为什么一些微生物中有这一系统,而其它的则不呢?也许CRISPR还有其它未知的基本生物学 功能?
“媒体大多将注意力放在了作为一种技术的CRISPR系统,当然这有很好的理由,这也是我们发现的令人难以置信的作用和机会。但是与此同时,还有很多有趣的基础生物学研究。”加州大学伯克利分校生物学家Jennifer Doudna说,她是首位将CRISPR-Cas作为一种基因编辑工具的科学家之一。
No.1 CRISPR从哪里来?
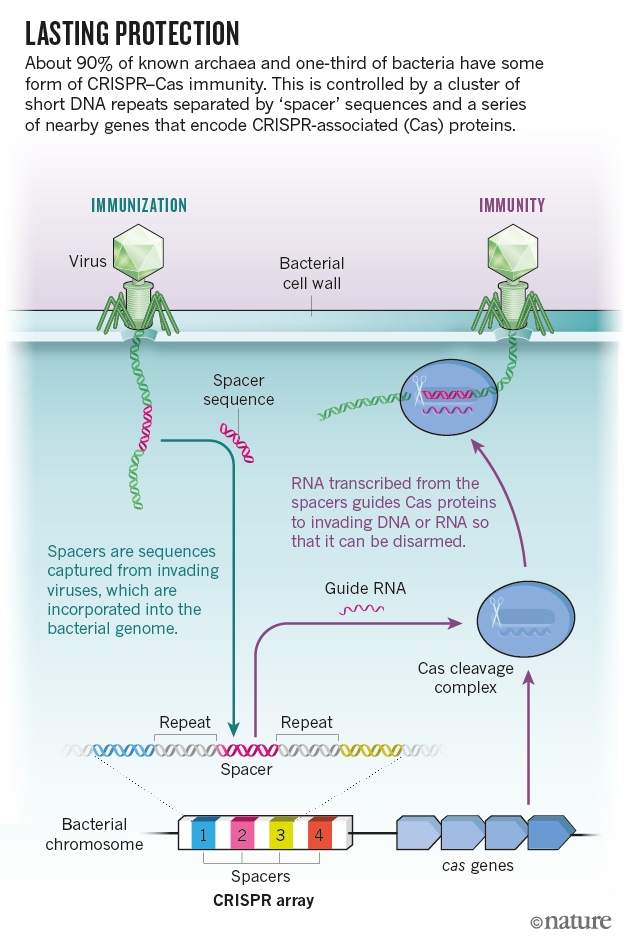
CRISPR-Cas的生物学优点已经很清楚了。原核生物,包括细菌和古细菌(一类不太知名的单细胞生物,其中许多生活在极端环境中)都需要面对遗传侵略者的不断攻击。病毒的数量可能比原核生物多出十倍,据说每两天能杀死全世界一半的细菌。原核生物还会交换被称为质粒的DNA碎片,其中包括寄生生物,它们从宿主中获取资源,如果宿主试图驱逐它们,机体分子就会迫使其自毁。这世界上似乎没有什么地方是安全的,从土壤到海洋,再到地球上最荒凉最不适合生存的地方,遗传入侵者们一直都存在。
原核生物在进化过程中已经发展出了许多武器来应对这些威胁。例如,在特定识别序列处或附近用于切割DNA的酶。但这些防御 并不灵敏,每种酶被需要编程才能识别某些序列,而且只有当微生物具有正确基因的拷贝时才受到保护。CRISPR-Cas动态性更强,这类似于人体抗体感染后 发展出的长期免疫方式,能进化和记住特殊的遗传入侵者。
荷兰瓦赫宁根大学的微生物学家John van der Oost说:“我们首次听说这个假说时,认为这对于简单的原核生物来说太复杂了。”
Mojica等人从位于与病毒基因组有时匹配的CRISPR回文重复之间的DNA序列中,推导出CRISPR-Cas的功能。从那时起,研究人员就发现在细菌和古细菌暴露于特定的病毒或质粒后,某些CRISPR相关(Cas)蛋白会将这些间隔序列添加到基因组中。由这些间隔序列生成的RNA能指导其它Cas蛋白切断所有匹配序列的入侵DNA或RNA。
那么细菌和古细菌如何出现这种复杂的免疫系统的呢?
这个问题还没有答案,但多数人认可的是,这个系统来自转座子,也就是可以从基因组中的一个位置跳到另一个位置的“跳跃基因”。美国国家卫生研究院的进化生物学家Eugene Koonin在马里兰州贝塞斯达和他的同事发现了一类编码蛋白Cas1的可移动遗传元件,Cas1参与间隔序列插入基因组的过程。这些“casposons(转座子元件,生物通注)”可能就是CRISPR-Cas免疫的起源。研究人员现在正在努力了解这些DNA如何从一个地方跳到另一个地方,之后他们还会追踪这种机制如何演化成复杂的CRISPR-Cas机制的。
No.2 CRISPR如何发挥作用的?
近年来的一些研究已经解释了关于Cas蛋白如何添加到间隔序列中的许多分子细节,但是病毒DNA在化学上几乎与宿主DNA相同。那么在包装有DNA的细胞中,蛋白如何指导是哪些DNA添加到了CRISPR-Cas“记忆”中?
这种风险很高:如果一种细菌不小心添加了一种自己的DNA,它可能会通过自身免疫自杀。正如立陶宛维尔纽斯大学的生物化学家Virginijus Siksnys所说的:“这些酶是一把双刃剑。”
来自北卡罗来纳州立大学的微生物学家 Rodolphe Barrangou认为,这可能是因为细菌和古细菌的庞大群体能承受一些错误:一些细胞自杀可能无关紧要,只要其它细胞可以在病毒攻击后继续茁壮成长就行。
事实上,当病毒入侵一个细菌生态系统时,通常1000万个细菌中只有大约一个细菌能正确获得一个间隔序列用以自卫。可能性 这么低,因此针对驱动间隔序列的因素进行研究很困难。
同样,要了解为什么只有这个细胞能够成功也很困难。“在实际发生时很难捕获这个‘成功的’细菌,”洛克菲勒大学微生物学家Luciano Marraffini解释道。
寻找确定合适间隔序列的方法,并提高它们的整合速率,将会有助于解开这个谜题。一些研究表明包含有CRISPR-Cas机制的细胞或许将其作为一种记录装置,对它们所遇到的DNA和RNA序列进行分类。这也许同样也能让研究人员追踪一段时间内接触环境化学物质后细胞的基因表达。
此外研究人员还希望能了解如何修剪旧的记忆,大多数带有CRISPR-Cas系统的微生物都含有几十个间隔序列,也有的只有一个。相反,古细菌Sulfolobus tokodaii则拥有5个CRISPR-Cas系统,包括458个间隔序列,占用了其基因组的1%之多。看起来好像没有必要一直保留旧的间隔Spacer,因为如果病毒通过突变以逃避CRISPR-Cas,那么间隔序列就会过时。而且保留额外的DNA可能会成为微生物的负担。“一种细菌不能无限制地扩张其基因组,”以色列魏茨曼科学研究所遗传学家Rotem Sorek说。
No.3 CRISPR还有其它的作用吗?
一些间隔序列的起源带来新的问题:目前观察到的间隔序列中能与DNA数据库中任何已知序列匹配的不超过3%。
这也就是说我们对病毒可能还知之甚少,大多数测序工作都集中在那些会感染人,牲畜或作物的病毒上。佐治亚大学的RNA生物学家Michael Terns说:“我们对细菌的敌人,特别是古生菌的敌人知之甚少。”
还有一种可能是,一些间隔序列是病毒的“鬼魂”,也就是已经不存在的,或者经过突变“认不出了”的序列。第三种可能性则越来越引发关注,研究人员也已经发现一些例子:CRISPR-Cas系统的功能并不限于防范遗传入侵者。在一些细菌中,CRISPR-Cas组分能调控DNA修复,基因表达和生物膜的形成。它们还可以决定细菌感染他人的能力,比如说引起军团病(Legionnaires' disease)的Legionella pneumophila必须具有Cas蛋白Cas2,才能感染其天然宿主变形虫。
“一个主要的问题是:除了防御外敌它还具有多少生物学功能?”麻省大学医学院的分子生物学家Erik Sontheimer指出,“这是未来几年的关注焦点。”
Sontheimer还补充说,CRISPR也有可能成为另一个类似RNA干扰的发现,RNA干扰是一种在植物,动物和 其它非原核生物中沉默基因表达的系统,RNA干扰在早期也被认为是防御机制,只是到后来研究人员才注意到其在调节宿主基因表达中的重要作用。
这也可以解释为什么一些间隔区不匹配已知的病毒或质粒,荷兰代尔夫特理工大学的微生物学家Stan Brouns说:“这些系统并不完美,它们捕获病毒DNA同时也捕获自己的DNA。只要它们不死,一旦开始捕获新的DNA片段,就可以获得新的功能 。”
No.4 为什么只有一些微生物采用这一系统?
无论CRISPR-Cas具有什么其它功能,显然只有一些微生物采用这一系统。超过90%的古细菌具有基于CRISPR的免疫系统,但只有约三分之一的已测序细菌出现了这一系统。所有的非原核生物,甚至单细胞生物,都没有发现有CRISPR-Cas。
一种被称为Nanoarchaeum equitans的古细菌作为寄生生物,寄生于另一个生活在接近沸点的水中的古细菌上,其基因组已经删去了与能量产生和一般细胞管理相关的许多基因。然而在其微小的只有490,000个碱基的DNA中,N. equitans还保留了约30个间隔序列的CRISPR-Cas系统。“一大部分的基因组仍然放在CRISPR上,”英国圣安德鲁斯大学的分子生物学家Malcolm White说:“CRISPR一定具有什么重要作用,但我们竟不知道其中的原因。”
英国埃克塞特大学微生物学家Edze Westra认为,这种差异表明一定存在有某些倾向于CRISPR-Cas系统的关键生态因素,使得病毒防御或其它益处比细胞自杀的风险更为重要。极端环境似乎有利于CRISPR-Cas系统,但Westra也指出, 在更舒适环境的细菌中,这种系统的出现频率也不相同。比如当一种禽类的病原体(Mycoplasma gallisepticum)的宿主从鸡转换为野生雀类时,这种细菌就丢弃了其CRISPR-Cas设备。为什么这个系统在鸡中就很有用,而在雀类中就不好用, 目前还不清楚。
数学模型和一些早期的实验室实验表明,当只有几种类型的病毒竞争时,CRISPR-Cas可能更有优势。在添加的DNA序列还没有成为基因组负荷时,CRISPR-Cas间隔序列可以记录有限数量的病毒序列。如果环境中的病毒多样性大大超过可能的间隔序列数量,那么CRISPR-Cas系统可能没有什么用处。
另一种可能性是在极端环境中的古生菌不能过度依赖其它防御手段。细菌阻止入侵者的一种常见方式是让自身外壳中称为包膜的蛋白质发生突变。然而,衣壳蛋白的结构对于在恶劣条件下生物的生存至关重要,因此一些古生菌可能难于修改这些衣壳蛋白。
No.5 有多少种CRISPR-Cas?
多数人倾向于CRISPR-Cas9系统,因为它可用于基因组编辑中,并且简单且具有多样性,但是微生物不喜欢。相反,他们倾向于混合和匹配不同的系统,迅速从其它细菌中学习新的细胞并丢弃旧的细胞。
研究人员已正式确认了6种不同类型的CRISPR系统,其中又有19种亚型。“我们其实只知道它们中的一小部分如何发挥作用的。”Marraffini说。
揭开这些机制可能是找到CRISPR-Cas系统新的生物技术应用的关键。例如,最受欢迎的CRISPR-Cas9是II型系统,它采用的是从间隔序列转录的RNA分子指导酶切割入侵的病毒或质粒DNA。但是在去年6月发现的VI型系统中的酶切割的是RNA,而不是DNA。另外IV型系统包含一些与CRISPR-Cas相关的基因,但缺乏重复和插入间隔序列的机制。
III型系统是自然界中最常见的CRISPR-Cas系统之一,但却是研究人员了解最少的一类系统。迄今为止的证据表明,它们响应的不是入侵的DNA或RNA本身,而是响应将DNA转录成RNA的过程。如果证明是这种情况,这将是一种新的调控形式,可以扩展基因组编辑的CRISPR-Cas工具箱。
此外,在研究人员将其搜索范围从培养的微生物扩展到来自环境DNA样品的遗传序列时,也有可能会突然出现其它系统。
对于Mojica来说,探索这种多样性,解答关于CRISPR系统的基本问题,比研究它们引发的技术革命更有趣。这让他的许多同事感到困惑,他说。Mojica沉浸在CRISPR-Cas生物学中已经25年了,虽然有很多资金涌向编辑基因组研究的科研人员,但是投入到他所做的工作的资金却很少。
Mojica说:“我知道这是一个伟大的工具。梦幻般的可用于治疗疾病,但这不是我的事。我想知道系统如何开始,又是如何结束的”。
参考资料: